Нуклеиновые кислоты
- Состав нуклеиновых кислот
- ДНК
- РНК
- Макромолекулярная структура ДНК
- Выделение дезоксирибонуклеиновых кислот
- Выделение рибонуклеиновых кислот
- Природа межнуклеотидных связей
- ДНК
- РНК
- Нуклеиновые кислоты, их значение
- Список литературы
Нуклеиновые кислоты представляют собой биополимеры. Их макромолекулы состоят из не однократно повторяющихся звеньев, которые представлены нуклеотидами. И их логично назвали полинуклеотидами. Одной из главных характеристик нуклеиновых кислот является их нуклеотидный состав. В состав нуклеотида (структурного звена нуклеиновых кислот) входят три составные части:
- азотистое основание. Может быть пиримидиновое и пуриновое. В нуклеиновых кислотах содержатся основания 4-х разных видов: два из них относятся к классу пуринов и два – к классу пиримидинов. Азот, содержащийся в кольцах, придает молекулам основные свойства.
- остаток фосфорной кислоты. Нуклеиновые кислоты являются кислотами потому, что в их молекулах содержится фосфорная кислота.
- моносахарид - рибоза или 2-дезоксирибоза. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т.е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два вида нуклеиновых кислот – рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дизоксирибозу.
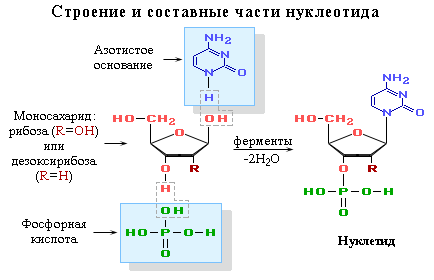
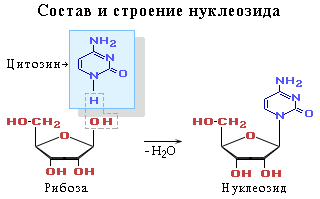
Нуклеотид по своей сути это фосфорный эфир нуклеозида. В состав нуклеозида входят два компонента: моносахарид (рибоза или дезоксирибоза) и азотистое основание.
На закате 40-х — начале 50-х годов, когда уже стали появляться такие методы исследования, как хроматография на бумаге и УФ-спектроскопия. Подтверждались многочисленные исследования нуклеотидного состава НК (Чаргафф, А. Н. Белозерский). Данные полученные при исследованиях в конец разрушили отжившие и некомпетентные представления о нуклеиновых кислотах, как о полимерах, содержащих повторяющиеся тетрануклеотидные последовательности — тетрануклеотидная теория строения ПК, господствовавшая в 30—40-е годы. Также дали основания для создания современных представлений не только о первичной структуре ДНК и РНК, но и об их макромолекулярной структуре и функциях.
Метод определения состава ПК основан на анализе гидролизатов, образующихся при их ферментативном или химическом расщеплении. Обычно используются три способа химического расщепления НК. Кислотный гидролиз в жестких условиях (70%-ная хлорная кислота, 100° С, 1 ч или 100%-ная муравьиная кислота, 175° C, 2 ч), применяемый для анализа как ДНК, так и РНК, приводит к разрыву всех N-гликозидных связей и образованию смеси пуриновых и пиримидиновых оснований. При исследовании РНК могут использоваться как мягкий кислотный гидролиз (1 н. соляная кислота, lOO° C, 1 ч), в результате которого образуются пуриновые основания и пирамидиповые нуклеозид-2'(3')-фосфаты, так и щелочной гидролиз (0,3 н. едкий кали, 37° С, 20 ч), дающий смесь нуклеозид -2' (3') -фосфатов.
Поскольку в НК число нуклеотидов каждого вида равно числу соответствующих оснований, для установления нуклеотидного состава данной НК достаточно определить количественное соотношение оснований. Для этой цели из гидролизатов с помощью хроматографии на бумаге или электрофореза (когда в результате гидролиза получают нуклеотиды) выделяют индивидуальные соединения. Каждое основание независимо от того, связано оно с углеводным фрагментом или нет, обладает характерным максимумом поглощения в УФ, интенсивность которого зависит от концентрации. По этой причине, исходя из УФ-спектров выделенных соединений, можно определить количественное соотношение оснований, а следовательно, и нуклеотидный состав исходной НК.
При количественном определении минорных нуклеотидов, особенно таких неустойчивых, как дигидроуридиловая кислота, пользуются ферментативными методами гидролиза (ФДЭ змеиного яда и селезенки).
Использование описанных выше аналитических приемов показало, что ПК различного происхождения состоят за редким исключением из четырех основных нуклеотидов и что содержание минорных нуклеотидов может меняться в значительных пределах.
Как будет показано далее, при изучении нуклеотидного состава ДНК были получены данные, которые помогли установить ее пространственную структуру.
При исследовании нуклеотидного состава нативных ДНК различного происхождения Чаргаффом были обнаружены следующие закономерности.
1. Все ДНК независимо от их происхождения содержат одинаковое число пуриновых и пиримидиновых оснований. Следовательно, в любой ДНК на каждый пуриновый нуклеотид приходится один пиримидиновый.
2. Любая ДНК всегда содержит в равных количествах попарно аденин и тимин, гуанин и цитозин, что обычно обозначают как А=Т и G=C. Из этих закономерностей вытекает третья.
3. Количество оснований, содержащих аминогруппы в положении 4 пиримидинового ядра и 6 пуринового (цитозин и аденин), равно количеству оснований, содержащих оксо-группу в тех же положениях (гуанин и тимин), т. е. A+C=G+T. Эти закономерности получили название правил Чаргаффа. Наряду с этим было установлено, что для каждого типа ДНК суммарное содержание гуанина и цитозина не равно суммарному содержанию аденина и тимина, т. е. что (G+C)/(A+T), как правило, отличается от единицы (может быть как больше, так и меньше ее). По этому признаку различают два основных типа ДНК: А Т-тип с преимущественным содержанием аденина и тимина и G C-тип с преимущественным содержанием гуанина и цитозина.
Величину отношения содержания суммы гуанина и цитозина к сумме содержания аденина и тимина, характеризующую нуклеотидный состав данного вида ДНК, принято называть коэффициентом специфичности. Каждая ДНК имеет характерный коэффициент специфичности, который может изменяться в пределах от 0,3 до 2,8. При подсчете коэффициента специфичности учитывается содержание минорных Оснований, а также замены основных оснований их производными. Например, при подсчете коэффициента специфичности для ЭДНК зародышей пшеницы, в которой содержится 6% 5-метилцитозина, Последний входит в сумму содержания гуанина (22,7%) и цитозина (16,8%). Смысл правил Чаргаффа для ДНК стал понятным после установления ее пространственной структуры.
Первые сведения о нуклеотидном составе РНК относились к препаратам, представляющим собой смеси клеточных РНК (рибосомных, информационных и транспортных) и называемым обычно суммарной фракцией РНК. Правила Чаргаффа в этом случае не соблюдаются, хотя определенное соответствие между содержанием гуанина и цитозина, а также аденина и урацила все же имеет, место.
Данные, полученные в последние годы при анализе индивидуальных РНК, показывают, что и на них правила Чаргаффа не распространяются. Однако различия в содержании аденина и урацила, а также гуанина и цитозина для большинства РНК невелики и что, следовательно, тенденция к выполнению указанных правил все же наблюдается. Этот факт объясняется особенностями макроструктуры РНК.
Характерными структурными элементами некоторых РНК являются минорные основания. Соответствующие им нуклеотидные остатки обычно входят в состав транспортных и некоторых других РНК в очень небольших количествах, поэтому определение полного нуклеотидного состава таких РНК представляет собой иногда весьма сложную задачу.
2. Макромолекулярная структура ДНК
В 1953 г. Уотсон и Крик, опираясь на известные данные о конформации нуклеозидных остатков, о характере межнуклеотидной связи в ДНК и закономерности нуклеотидного состава ДНК (правила Чаргаффа), расшифровали рентгенограммы паракристаллической формы ДНК [так называемой В-формы, образующейся при влажности выше 80% и при высокой концентрации противоионов (Li+) в образце]. Согласно их модели, молекула ДНК представляет собой правильную спираль, образованную двумя полидезоксирибонуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Диаметр спирали практически постоянен вдоль всей ее длины и равен 1,8 нм (18 А).
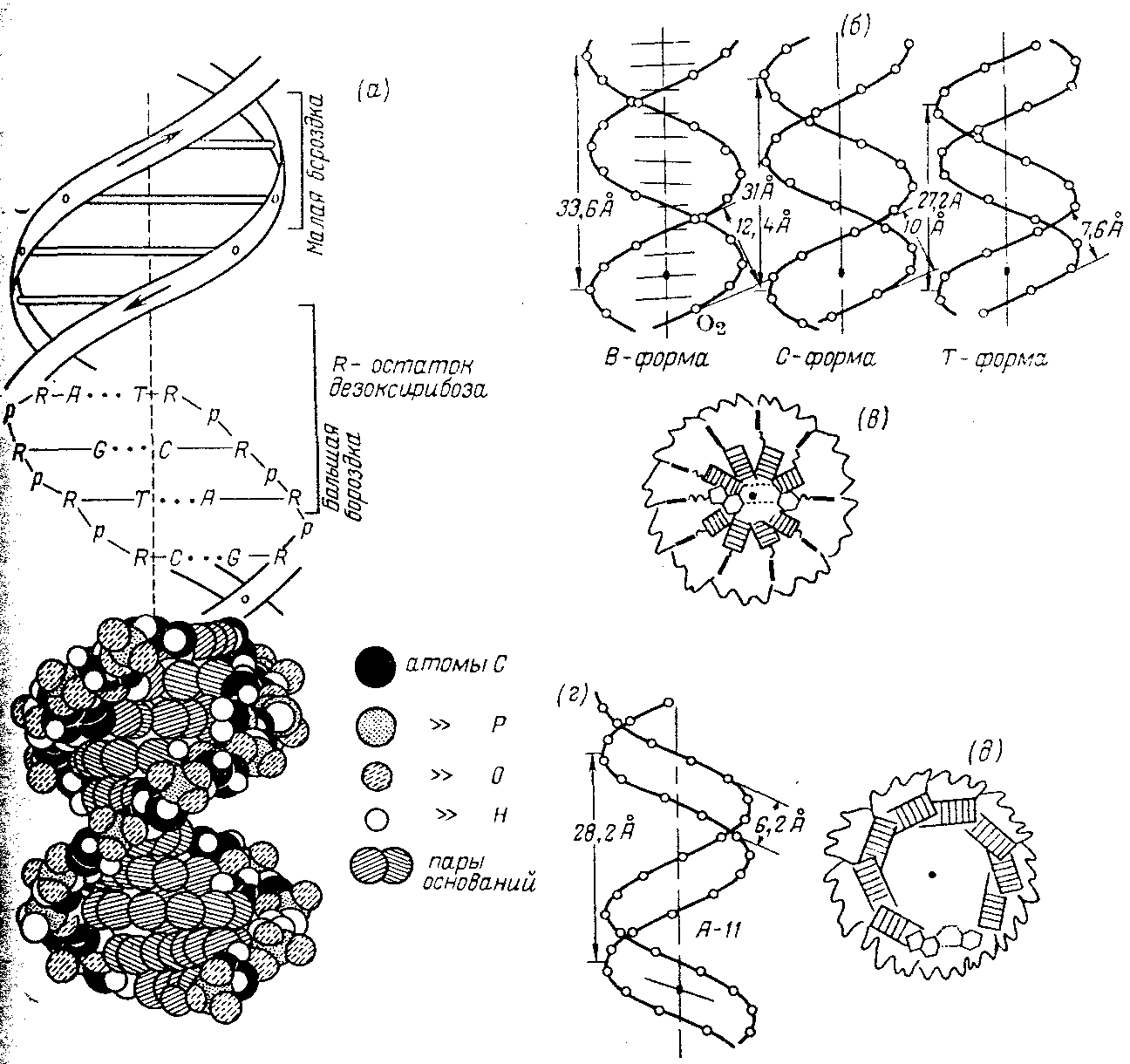
Макромолекулярная структура ДНК.
(а)—Модель Уотсона — Крика;
(6)—параметры спиралей В-, С- и Т-форм ДНК (проекции перпендикулярно оси спирали);
(в)—поперечный разрез спирали ДНК в В-форме (заштрихованные прямоугольники изображают пары оснований);
(г)—параметры спирали ДНК в А-форме;
(д)—поперечный разрез спирали ДНК в А-форме.
Длина витка спирали, который соответствует ее периоду идентичности, составляет 3,37 нм (33,7 А). На один виток спирали приходится 10 остатков оснований в одной цепи. Расстояние между плоскостями оснований равно, таким образом, примерно 0,34 нм (3,4 А). Плоскости остатков оснований перпендикулярны длинной оси спирали. Плоскости углеводных остатков несколько отклоняются от этой оси (первоначально Уотсон и .Крик предположили, что они параллельны ей).
Из рисунка видно, что углеводофосфатный остов молекулы обращен наружу. Спираль закручена таким образом, что на ее поверхности можно выделить две различные по размерам бороздки (их часто называют также желобками) — большую, шириной примерно 2,2 нм (22 А), и малую —шириной около 1,2 нм (12А). Спираль — правовращающая. Полидезоксирибонуклеотидные цепи в ней антипараллельны: это означает, что если мы будем двигаться вдоль длинной оси спирали от одного ее конца к другому, то в одной цепи мы будем проходить фосфодиэфирные связи в направлении 3'а 5', а в другой — в направлении 5'а 3'. Иными словами, на каждом из концов линейной молекулы ДНК расположены 5'-конец одной и 3'-конец другой цепи.
Регулярность спирали требует, чтобы против остатка пуринового основания в одной цепи находился остаток пиримидинового основания в другой цепи. Как уже подчеркивалось, это требование реализуется в виде принципа образования комплементарных пар оснований, т. е. остаткам аденина и гуанина в одной цепи соответствуют остатки тимина и цитозина в другой цепи (и наоборот).
Таким образом, последовательность нуклеотидов в одной цепи молекулы ДНК предопределяет нуклеотидную последовательность другой цепи.
Этот принцип является главным следствием модели Уотсона и Крика, поскольку он в удивительно простых химических терминах объясняет основное функциональное назначение ДНК — быть хранителем генетической информации.
Заканчивая рассмотрение модели Уотсона и Крика, остается добавить, что соседние пары остатков оснований в ДНК, находящейся в В-форме, повернуты друг относительно друга на 36° (угол между прямыми, соединяющими атомы С 1 ' в соседних комплементарных парах).
3. Выделение дезоксирибонуклеиновых кислот
Живые клетки, за исключением сперматозоидов, в норме содержат значительно больше рибонуклеиновой, чем дезоксирибонуклеиновой кислоты. На методы выделения дезоксирибонуклеиновых кислот оказало большое влияние то обстоятельство, что, тогда как рибонуклеопротеиды и рибонуклеиновые кислоты растворимы в разбавленном (0,15 М) растворе хлористого натрия, дезоксирибонуклеопротеидные комплексы фактически в нем нерастворимы. Поэтому гомогенизированный орган или организм тщательно промывают разбавленным солевым раствором, из остатка с помощью крепкого солевого раствора экстрагируют дезоксирибонуклеиновую кислоту, которую осаждают затем добавлением этанола. С другой стороны, элюирование того же остатка водой дает раствор, из которого при добавлении соли выпадает дезоксирибонуклеопротеид. Расщепление нуклеопротеида, который в основном представляет собой солеподобный комплекс между полиосновными и поликислотными электролитами, легко достигается растворением в крепком солевом растворе или обработкой тиоцианатом калия. Большую часть белка можно удалить либо добавлением этанола, либо эмульгированием с помощью хлороформа и амилового или октилового спирта (белок образует с хлороформом гель). Широко применялась также обработка детергентами. Позднее дезоксирибонуклеиновые кислоты выделяли с помощью экстракции водными n-аминосалицилат — фенольными растворами. При использовании этого метода были получены препараты дезоксирибонуклеиновой кислоты, из которых одни содержали остаточный белок, тогда как другие были фактически свободны от белка, что указывает на то, что характер связи белок — нуклеиновая кислота различен в различных тканях. Удобная модификация состоит в гомогенизировании животной ткани в 0,15 М растворе фенолфталеиндифосфата с последующим добавлением фенола для осаждения ДНК (свободной от РНК) с хорошим выходом.
Дезоксирибонуклеиновые кислоты, каким бы способом они не выделялись, представляют собой смеси полимеров различного молекулярного веса, за исключением образцов, полученных из некоторых видов бактериофагов.
ФРАКЦИОНИРОВАНИЕ
Ранний метод разделения заключался в фракционной диссоциации гелей дезоксирибонуклеопротеида (например, нуклеогистона) посредством экстракции водными растворами хлористого натрия увеличивающейся молярности. Таким путем препараты дезоксирибонуклеиновой кислоты были разделены на ряд фракций, характеризующихся различным отношением содержания аденина с тимином к сумме гуанина с цитозином, причем более легко выделялись фракции, обогащенные гуанином и цитозином. Сходные результаты были получены при хроматографическом отделении дезоксирибонуклеиновой кислоты от гистона, адсорбированного на кизельгуре, с применением градиентного элюирования растворами хлористого натрия. В улучшенном варианте этого метода очищенные фракции гистона сочетались с n-аминобензилцеллюлозой с образованием диазомостиков от тирозиновых и гистидиновых групп белка. Описано также фракционирование нуклеиновых кислот на метилированном сывороточном альбумине (с кизельгуром в качестве носителя). Скорость элюирования с колонки солевыми растворами увеличивающейся концентрации зависит от молекулярного веса, состава (нуклеиновые кислоты с высоким содержанием гуанина с цитозином элюируются легче) и вторичной структуры (денатурированная ДНК прочнее удерживается колонкой, чем нативная). Таким способом из ДНК морского краба Cancer borealis выделен природный компонент — полидезоксиадениловая-тимидиловая кислота. Фракционирование дезоксирибонуклеиновых кислот проводилось также посредством градиентного элюирования с колонки, наполненной фосфатом кальция.
4. Выделение рибонуклеиновых кислот
Методы, используемые для экстракции рибонуклеиновых кислот, частично зависят от природы органа или организма. В одном из ранних методов, использованном Левиным, к густому тесту из дрожжей добавляли щелочь, смесь перемешивали с пикриновой кислотой, фильтровали и нуклеиновую кислоту осаждали из фильтрата добавлением соляной кислоты. Такая довольно жесткая обработка приводила к тому, что полученная нуклеиновая кислота значительно отличалась от “нативной” рибонуклеиновой кислоты. Для выделения рибонуклеиновых кислот, приближающихся по структуре к нуклеиновым кислотам живой клетки, необходимо избегать применения жестких условий (рН, температура) в то же время необходимо, насколько возможно, затормозить ферментативный распад. Широко применялась экстракция рибонуклеопротеидов изотоническим раствором хлористого натрия. Белки от нуклеиновых кислот могут быть отщеплены различными методами, такими, как обработка смесями хлороформа с октиловым спиртом, додецилсульфатом натрия, нитратом стронция или спиртом, а также расщепление белковой фракции трипсином. И снова эффективность каждого метода определяется природой рибонуклеопротеида. Для инактивации ферментов в процессе экстракции полезно применение хлоргидрата гуанидина (денатурирующего агента); для выделения рибонуклеиновых кислот и нативных рибонуклеопротеидов из дрожжей был применен метод, использующий адсорбцию рибонуклеаз на бентоните после предварительной обработки ионами цинка.
Особые преимущества имеет выделение рибонуклеиновых кислот из гомогенатов тканей млекопитающих, микроорганизмов и вирусов экстракцией фенолом и водой при комнатной температуре, так как при этом белки и дезоксирибонуклеиновые кислоты выпадают в осадок, активность рибонуклеазы подавляется и высокополимерные продукты могут быть получены с хорошими выходами. Прямая экстракция дрожжей водным раствором фенола была применена для препаративного получения транспортных РНК.
ФРАКЦИОНИРОВАНИЕ
Помимо ряда вирусных нуклеиновых кислот, большинство выделенных полирибонуклеотидов, бесспорно, представляют собой сложные смеси, содержащие полимеры с различной длиной цепи, нуклеотидной последовательностью и составом оснований (присутствие или отсутствие “минорных” оснований). Существует ряд приемов для частичного фракционирования, однако, пока не разработаны удовлетворительные методы характеристики, трудно определить степень чистоты или гомогенности рибонуклеиновых кислот. В основу оценки чистоты транспортных РНК, этих сравнительно низкомолекулярных полирибонуклеотидов, может быть положена их ферментативная реакция с аминокислотами (через аминоациладенилаты), что, конечно, позволяет оценить и их биохимическую однородность.
Методы фракционирования включают осаждение нейтральными солями, электрофорез, хроматографию на фосфате кальция и осаждение дигидрострептомицином. Недавно для фракционирования рибонуклеиновых кислот была использована фракционная диссоциация комплексов нуклеиновая кислота — гистон, примененная ранее к дезоксинуклеиновым кислотам. Во всех фракциях отношение 6-амино- к 6-кетонуклеозидам было близко к единице. В некоторой степени фракционирование происходит при экстракции фенолом, возможно как результат дифференциального связывания нуклеиновых кислот с белками. Анионообменные целлюлозы, такие как ЭКТЕОЛА и ДЭАЭ, широко применяются в настоящее время для фракционирования не только рибонуклеиновых кислот, включая специфичные для аминокислот транспортные РНК, но и рибонуклеопротеидов и даже вирусных препаратов. Для элюирования обычно используют растворы нейтральных или близких к нейтральным солеи. Поразительной особенностью метода является способность этих ионообменников к разделению очень широкого спектра веществ, начиная от изомеров мононуклеотидов и олигонуклеотидов с различной длиной цепи или различного состава и кончая полинуклеотидами чрезвычайно высокого молекулярного веса. Опубликовано сообщение о разделении на колонках из ДЭАЭ-декстрана РНК, меченной валином, от немеченой акцепторной РНК. Для фракционирования рибонуклеиновых кислот были также применены модифицированные ионообменные целлюлозы, в которых к целлюлозе с помощью эпихлоргидрина присоединены нуклеозиды (вместо триэтаноламина), особенно аденозин и гуанозин [44]. Подобное использование ЭКТЕОЛА-целлюлозы для фракционирования или выделения информационной РНК, связанной в данный момент с ДНК, основано на способности к специфическому образованию водородных связей: ЭКТЕОЛА связывает денатурированную ДНК данного организма (для элюирования ДНК необходим растворитель чрезвычайно высокой ионной силы), а информационная РНК элюируется растворами понижающейся ионной силы. Посредством хроматографии на трет-аминоалкилированном крахмале транспортная рибонуклеиновая кислота была разделена на фракции на основании повышенного сродства к тирозину и лейцину. Хроматография на оксиапатите дает хорошее разделение рибонуклеиновых кислот, специфичных для валина и фенилаланина.
В другом методе, имеющем значительную потенциальную ценность, используется поперечно сшитый полидиазостирол, полученный в результате реакции полиаминостирола с азотистой кислотой; метод основан на наблюдениях, что соединения диазония легко реагируют с некоторыми аминокислотами с образованием ковалентно связанных производных. В пределах рН от 7 до 8,5 быстро реагируют только тирозин и гистидин. Препараты транспортных РНК, полностью этерифицированные аминокислотами, встряхивали с нерастворимым полидиазостиролом, который реагировал только с нуклеиновыми кислотами, меченными тирозином и гистидином.
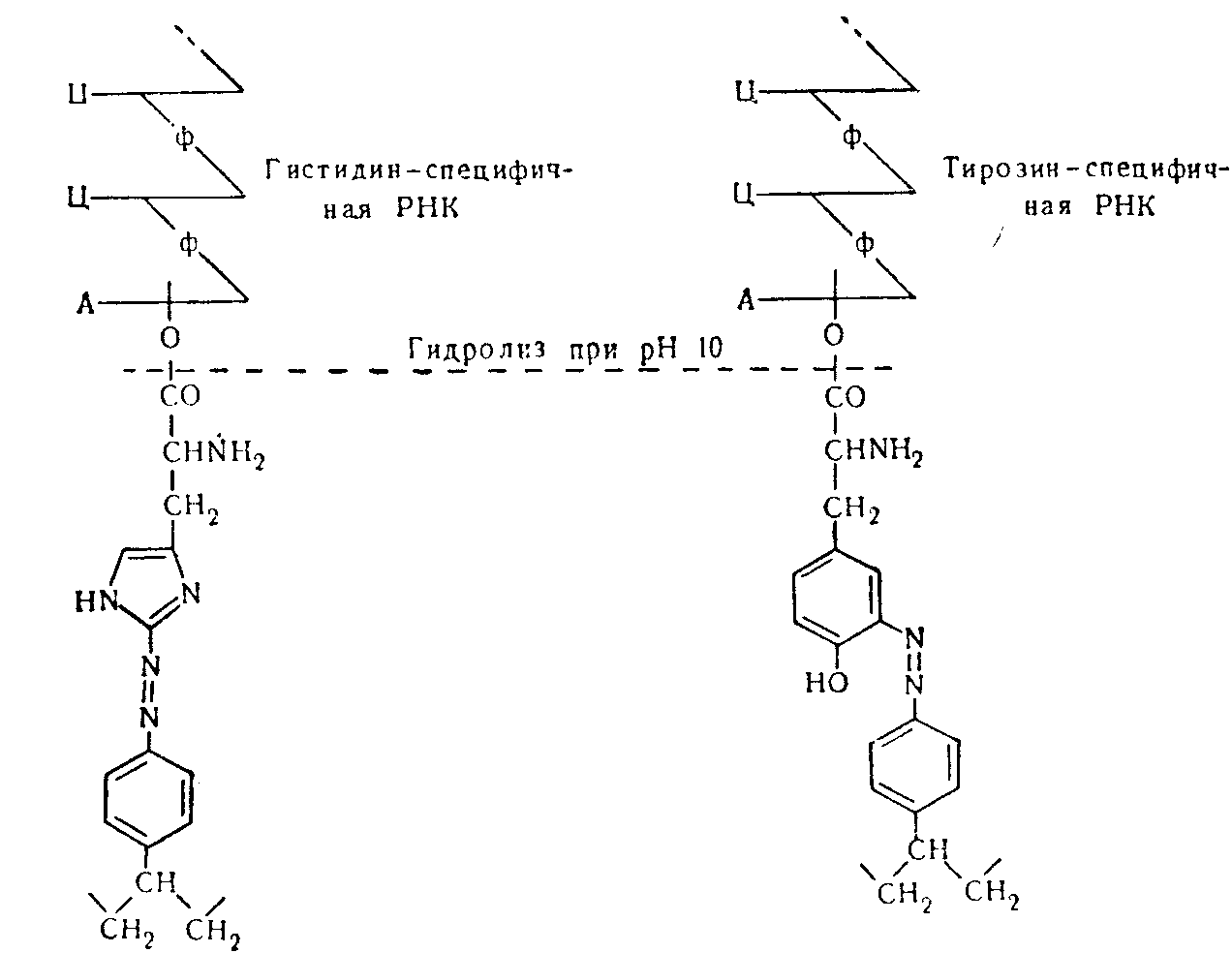
Дальнейшая очистка достигалась повторной этерификацией тирозином при использовании очищенного тирозин-активнрующего фермента и повторной обработкой полидиазостиролом. С неэтерифицированной специфичной к гистидину рибонуклеиновой кислотой реакции не происходило, и она оставалась в растворе, в то время как специфичная к тирозину нуклеиновая кислота освобождалась, как и прежде, при обработке щелочью в мягких условиях. Обе фракции получены почти чистыми в отношении их аминокислотоакцепторной специфичности. Предварительные наблюдения показали, что специфичная к валину рибонуклеиновая кислота, вполне вероятно, может быть этерифицирована дипептидом тирозилвалином.
5. Природа межнуклеотидных связей
Работы по определению способа соединения нуклеотидов в полимерных молекулах НК были успешно завершены в начале 50-х годов сразу после того, как была установлена структура нуклеотидов и изучены некоторые свойства их производных (главным образом эфиров). К этому же времени были разработаны методы выделения и очистки ДНК и РНК, так что исследование природы межмономерных связей проводилось с использованием чистых, хотя и сильно деградированных препаратов НК.
Первые сведения о типе межмономерной, или, как ее принято называть, межнуклеотидной связи были получены с помощью потенциометрического титрования. Эти сведения свидетельствовали о наличии как в РНК, так и в ДНК только одной гпдроксильной группы у каждой фосфатной группы (рКа~1). На основании этого было сделано заключение, что НК содержит структурную единицу дизамещенной фосфорной кислоты.
Естественно было предположить, что фосфатные остатки “сшивают” нуклеозиды за счет двух своих гидроксилов, а один остается свободным. Оставалось выяснить, какие части нуклеозидных фрагментов участвуют в образовании связи с фосфатными группами.
Поскольку НК могут быть дезаминированы действием азотистой кислоты, очевидно, что аминогруппы пиримидиновых и пуриновых оснований не принимают участия в образовании межнуклеотидной связи. Помимо этого потенциометрическое титрование указывало, что и оксо(окси)-группы остатков гуанина и урацила, входящих в состав НК, свободны. На основании этих данных было сделано заключение о том, что межнуклеотидные связи образованы фосфатной группой и гидроксильными группами углеводных остатков (т. е. что они являются фосфодиэфирными), которые, следовательно, и являются ответственными за образование полимерной цепи (НК). Таким образом, то, что принято обычно называть межнуклеотидной связью, представляет собой по существу узел, включающий систему связей:
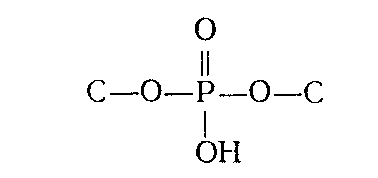
(где С — первичный или вторичный атомы углерода остатка углевода). При гидролизе ДНК и РНК в зависимости от условий реакции, образуются нуклеотиды с разным положением фосфатного остатка:
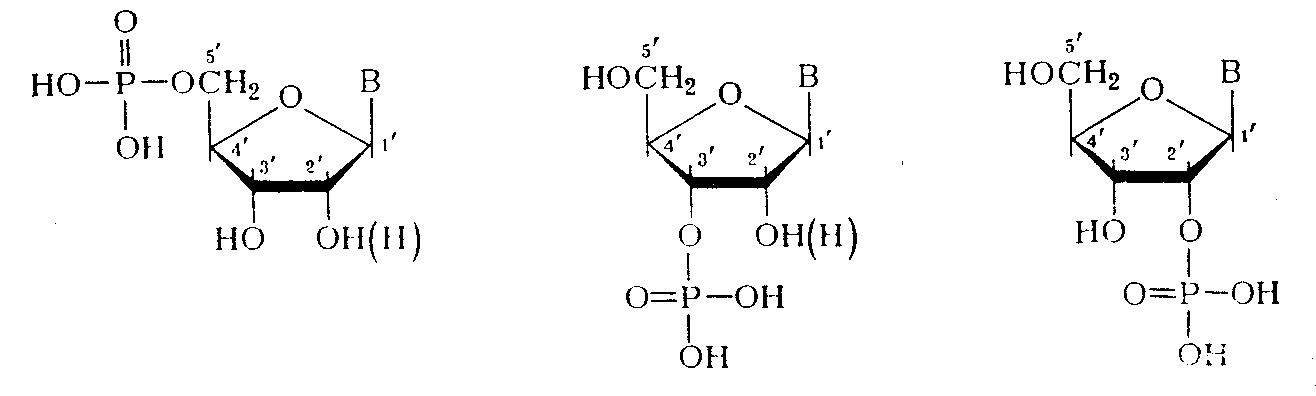
Если предположить, что в НК все межнуклеотидные связи идентичны, то, очевидно, что они могут включать помимо фосфатного остатка только З'-гидроксильную группу одного нуклеозидного звена и 5'-гидроксильную группу другого нуклеозидного звена (3'—У-связь). В случае же их неравноценности в полимерной цепи ДНК могли бы одновременно существовать три типа связей: 3'—5', 3'—3' и 5'—5'. Для РНК за счет участия 2 / -гидpoкcилы I-oй группы число типов связи должно было быть еще больше.
Установить истинную природу межнуклеотидных связей в нативных ДНК и РНК удалось в результате направленного расщепления биополимеров с помощью химического и ферментативного гидролиза и последующего выделения и идентификации полученных при этом фрагментов.
Химический гидролиз ДНК как метод деградации полимера с целью установления природы межнуклеотидной связи оказался практически непригодным. ДНК не расщепляется при щелочных значениях рН, что хорошо согласуется с предположением о фосфодиэфирной природе межнуклеотидной связи (устойчивость диалкилфосфатов в щелочной среде обсуждалась в разделе). При обработке кислотой даже в мягких условиях ДНК расщепляется как по фосфодиэфирным, так и по N-гликозидным связям, образованным пуриновыми основаниями. Вследствие этого расщепление полимера протекает неоднозначно, но из продуктов кислотного гидролиза ДНК все же удалось выделить ди-фосфаты пиримидиновых дезоксинуклеозидов, которые оказались идентичными синтетическим 3',5'-дифосфатам дезоксицитидина и дезокситимидина:
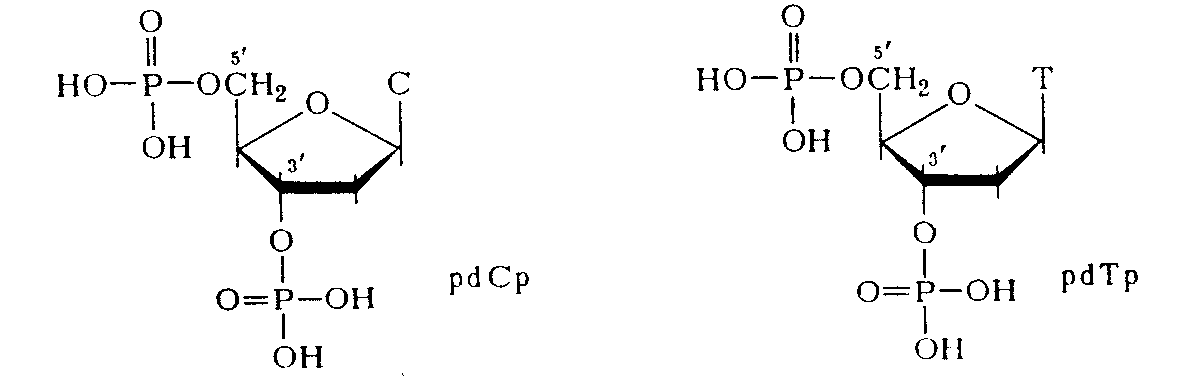
Здесь же важно отметить, что наличие этих соединений в продуктах деградации ДНК указывает на участие обеих гидроксильных групп, по крайней мере пиримидиновых мономерных компонентов, в образовании межнуклеотидной связи.
Более специфическим оказалось ферментативное расщепление ДНК. При обработке препаратов ДНК фосфодиэстеразой (ФДЭ) змеиного яда полимер практически полностью гидролизуется до дезоксннуклеозид-5'-фосфатов, структура которых была Остановлена сравнением с соответствующими нуклеотидами, полученными встречным синтезом.
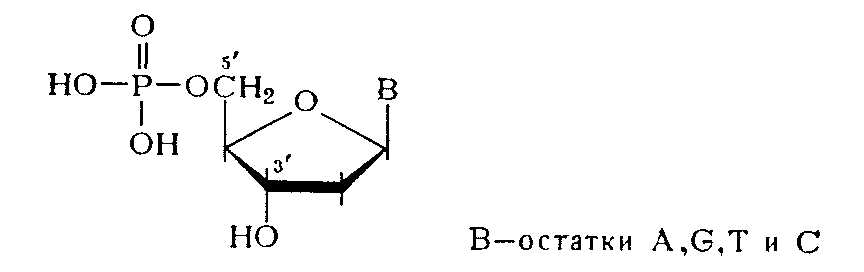
Эти данные свидетельствуют об участии 5'-гидроксильных групп всех четырех дезоксинуклеозидов, входящих в состав ДНК, в образовании межнуклеотидной связи. Аналогично, но до 3'-фосфатов дезоксинуклеозидов расщепляется ДНК в присутствии ФДЭ, выделенной из Микрококков или из селезенки.
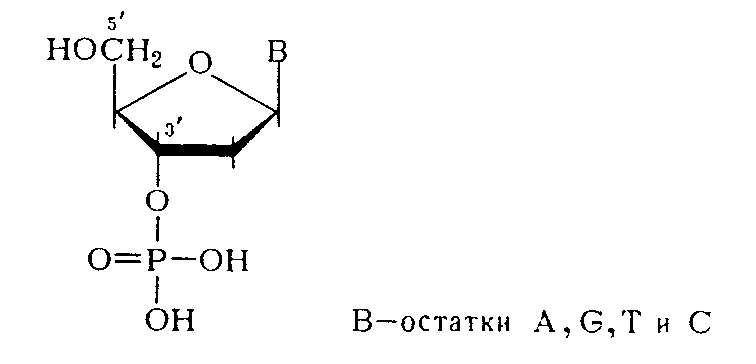
Из данных гидролиза ДНК фосфодиэстеразами различной специфичности становится очевидным, что связь нуклеозидных остатков в ДНК осуществляется фосфатной группой, которая одновременно этерифицирует гидроксильную группу у вторичного атома углерода (положение 3') одного нуклеозидного звена и гидроксильную группу у первичного атома углерода (положение 5') - другого нуклеотидного звена.
Таким образом, было убедительно доказано, что в ДНК межнуклеотидная связь осуществляется за счет фосфатной группы, а также 3'- и 5'-гидроксильных групп нуклеозидных остатков [(а) и (б) — направления расщепления полинуклеотидной цепи ДНК фосфодиэстеразами соответственно змеиного яда и селезенки или микрококков]:
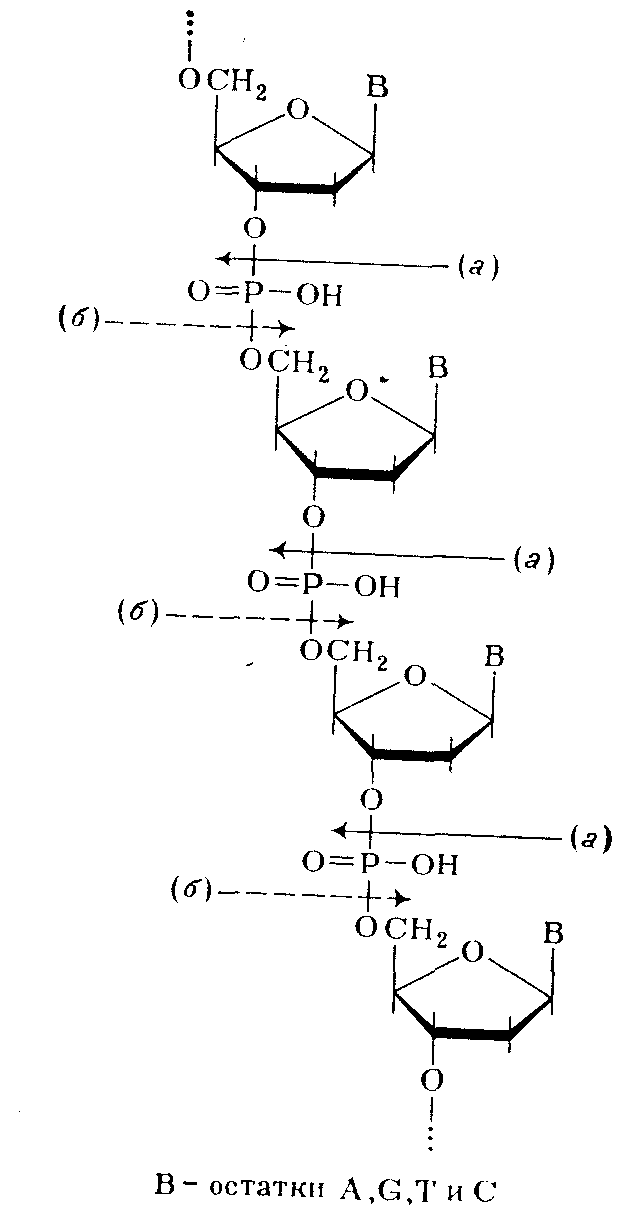
Предположение о возможности иного строения полимера с регулярно перемежающимися связями нуклеозидных остатков по типу 3'—3' и 5'—5' было отвергнуто, так как оно не удовлетворяло всем экспериментальным данным. Так, полимер такого типа не должен был бы полностью гидролизоваться (до мономеров) в присутствии ФДЭ змеиного яда, избирательно расщепляющей только алкиловые эфиры нуклеозид-5' –фосфатов. То же можно сказать о ФДЭ селезенки, селективно гидролизирующей алкиловые эфиры нуклеозид-3'-фосфатов.
Самым неясным и сложным оказался вопрос о природе межнуклеотидной связи в РНК. Уже на начальных стадиях при изучении строения РНК было установлено что, они чрезвычайно неустойчивы при щелочном гидролизе. Основными продуктами щелочного гидролиза РНК являются рибонуклсозид-2'- и рибонуклеозид-З'-фосфаты, образующиеся практически в равных количествах.
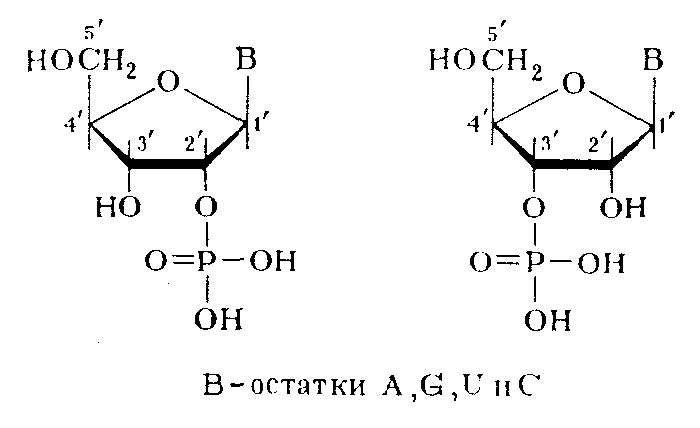
Рибонуклеозид-5'-фосфаты при этом не образуются. Эти данные не укладывались в представления о фосфодиэфирной природе межнуклеотидной связи в РНК и требовали всестороннего изучения. Очень важную роль в таком исследовании, которое выполнили в начале 50-х гг. Тодд с сотрудниками, сыграли синтетические алкиловые эфиры рибонуклеотидов, которые были получены специально, чтобы промоделировать тот или иной тип фосфодиэфирной связи.
Исследования школы Тодда предоставили данные о механизмах превращения алкиловых эфиров рибонуклеотидов в щелочной среде позволили предположить, что в РНК, так же как и в ДНК, межнуклеотидная связь осуществляется фосфатной группой и 3'- и 5'-гидроксильными группами углеводных остатков. Подобная связь в РНК должна очень легко расщепляться в щелочной среде, так как соседняя 2'-гидроксильная группа должна катализировать этот процесс при рН>10, когда начинается ионизация гидроксильных групп рибозы. Очень важно подчеркнуть, что промежуточными соединениями при щелочном расщеплении должны быть все четыре рибонуклеозид-2',З'-циклофосфата, а конечными — образующиеся при их гидролизе рибонуклеозид-3'-фосфаты и рибонуклеозид-2'-фосфаты (четыре пары изомеров).
Данные щелочного гидролиза ограничили количество возможных для РНК типов межнуклеотидных связей, но не прояснили вопроса о том, как построен этот полимер.
Самые точные сведения о типе межнуклеотидной связи в РНК, как и в случае ДНК, были получены с помощью ферментативного гидролиза.
Гидролиз РНК с использованием ФДЭ змеиного яда, протекающий до рибонуклеозид-5'-фосфатов, подтвердил уже прямым путем предположение об участии 5'-гидроксильных групп в образовании фосфодиэфирной связи между мономерными звеньями.
В дальнейшем были получены данные, на основе которых можно утверждать, что это действительно так (в результате открытия фосфоролиза РНК в присутствии фермента полинуклеотидфосфорилазы (ПНФаза), приводящего к образованию рибонуклеозид-5'-пирофосфатов):
![]()
Надо было только выяснить природу второй гидроксильной группы, участвующей в образовании межнуклеотидной связи. Частично решить эту задачу помог еще один фермент, который использовался для направленного расщепления РНК, — пиримидиловая рибонуклеаза (РНаза).
Ранее было показано, что этот фермент расщепляет только алкиловые эфиры пиримидиновых рибонуклеозид-3'-фосфатов до рибонук-леозид-3'-фосфатов (через промежуточный рибонуклеозид-2',З'-циклофосфат). Оказалось, что аналогичным образом этот фермент действует и на РНК. В экспериментах с любыми образцами очищенной РНК было обнаружено, что количество фосфорной кислоты, которая образуется при обработке полимера последовательно пиримидиловой РНазой и фосфомоноэстеразой (ФМЭ), а также количество иодной кислоты, затрачиваемой на последующее окисление, эквивалентно количеству остатков пиримидинов в данном образце РНК. Это говорило в пользу того, что по крайней мере пиримидиновые нуклеотиды в РНК связаны с соседними нуклеотидами только посредством 3'—5'-межнук-леотидной связи. Этот вывод подтверждают данные щелочной обработки ферментативных гидролизатов РНК, полученных после действия на нее РНазы: в щелочной среде миграция фосфатного остатка в рибонуклеозид-З'- и -2'-фосфатах невозможна, и наличие в соответствующих гидролизатах только пиримидиновых рибонуклеозид-З'-фосфатов делает очевидным 3'—5'-тип межнуклеотидной связи для пиримидиновых нуклеотидов.
6. Нуклеиновые кислоты, их значение
Значение нуклеиновых кислот очень велико. Некоторые особенности в химическом строения обеспечивают возможность ранения, переноса в цитоплазму и передачи по наследству дочерним клеткам информации о структуре белковых молекул, которые синтезируются в каждой клетке. Белки обусловливают большинство свойств и признаков клеток. Понятно поэтому, что стабильность структуры нуклеиновых кислот - важнейшее условие нормальной жизнедеятельности клеток и организма в целом. Любые изменения строения нуклеиновых кислот влекут за собой изменения структуры клеток или активности физиологических процессов в них, влияя таким образом на жизнеспособность.
Известно два типа нуклеиновых кислот: ДНК и РНК. ДНК (дезоксирибонуклеиновая кислота) - биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие одно из четырех азотистых оснований: аденин (А) или тимин (Т), цитозин (Ц) или гуанин (Г); пятиатомный сахар пентозу - дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов. В каждой цепи нуклеотиды соединяются путем образования ковалентных связей между дезоксирибозой одного и остатком фосфорной кислоты последующего нуклеотида. Объединяются две цепи в одну молекулу при помощи водородных связей, возникающих между азотистыми основаниями, входящими в состав нуклеотидов, образующих разные цепи. Количество таких связей между разными азотистыми основаниями неодинаково и вследствие этого они могут соединяться только попарно: азотистое основание А одной цепи полинуклеотидов всегда связано двумя водородными связями с Т другой цепи, а Г - тремя водородными связями азотистым основанием Ц противоположной полинуклеотидной цепочки. Такая способность к избирательному соединению нуклеотидов называется комплиментарностью. Комплиментарное взаимодействие нуклеотидов приводит к образованию пар нуклеотидов. В полинуклеотидной цепочке соседние нуклеотиды связаны между собой через сахар и остаток фосфорной кислоты.
РНК (рибонуклеиновая кислота), так же как ДНК, представляет собой полимер мономерами которого служат нуклеотиды. Азотистые основания те же самые, что входят в состав ДНК (аденин, гуанин, цетозин); четвертое - урацил - присутствует в молекуле РНК вместо тимина. Нуклеотиды РНК содержат вместо дизоксирибозы другую пентозу - рибозу. В цепочке РНК нуклеотиды соединяются путем образования ковалентных связей между рибозой одного нуклеотида и остатком фосфорной кислоты другого.
Известны двух- и одноцепочные молекулы рибонуклеиновой кислоты. Двухцепочные РНК служат для хранения и воспроизведения наследственной информации у некоторых вирусов, т.е. выполняют у них функции хромосом. Одноцепочные РНК осуществляют перенос информации о последовательности аминокислот в белках от хромосомы к месту их синтеза и участвуют в процессах синтеза.
Существует несколько видов одноцепочных РНК. Их названия обусловлены выполняемой функцией или местом нахождения в клетке. Основную часть РНК цитоплазмы (80-90%) составляет рибосомальная РНК (рРНК). Она содержится в органоидах клетки, осуществляющих синтез белков, - рибосомах. Размеры молекул рРНК относительно невелики, они содержат от 3 до 5 тысяч нуклеотидов. Другой вид РНК - информационные (иРНК), переносящие от хромосом к рибосомам информацию о последовательности аминокислот в белках, которые должны синтезироваться. Транспортные РНК (рРНК) включают 76-85 нуклеотидов и выполняют несколько функций. Они доставляют аминокислоты к месту синтеза белка, “узнают” (по принципу комплиментарности) участок иРНК, соответствующий переносимой аминокислоте, осуществляет аминокислоты на рибосоме.
- Н. Грин, У. Стаут, Д. Тейлор – Биология.
- З.А. Шабарова и А.А. богданов – Химия нуклеиновых кислот и их полимеров.
- А.П. Пехов – Биология и общая гинетика.
- 2. А. Микельсон – Химия нуклеозидов и нуклеотидов.
- З. Гауптман, Ю. Грефе, Х. Ремане – Органическая химия.
 При любом использовании материалов установите обратную ссылку на своем сайте.
При любом использовании материалов установите обратную ссылку на своем сайте.
